Takashi Suzuki's Lab.
Nagatsuta 4259 Suzukakedai-Campus,
Midoriku, Yokohama, JAPAN
TEL +81-(0)45-924-5796
FAX +81-(0)45-924-5974
Molecular mechanisms underlying axon targeting of photoreceptor cells
|
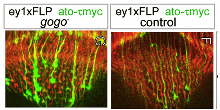
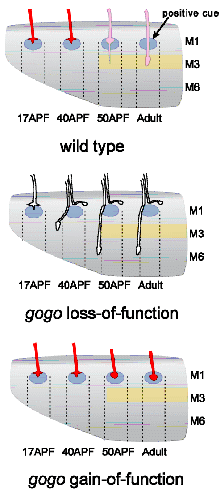
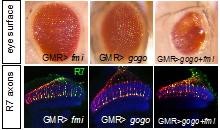
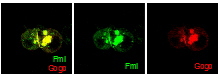
In a genetic screen, we identified a new axon guidance receptor, Golden Goal (Gogo), involved in different steps of R8 axon pathfinding (Tomasi et al., 2008). First, during larval development, Gogo mediates repulsive interactions between R8 axons, ensuring proper spacing of axons as they travel toward their target region, the medulla. As shown in Fig. 1.B, gogo mutant axons, stained in green, form bundles with each other, due to a loss of repulsive axon-axon interaction. In contrast, wild type axons are evenly spaced (Fig.1A). Later in development, during pupal stages, Gogo is involved in layer targeting of R8 axons. R8s target their synaptic layer in two steps (Fig. 2A): they first stop at the superficial layer of the medulla M1, and they reach their final layer M3 only later in development. To determine the role of Gogo in R8 axon targeting, we performed a detailed analysis of gogo mutant and overexpression phenotypes. We found that in gogo mutants, R8 axons often stray at the surface of the medulla and fail to enter medulla columns (Fig.2B). In Gogo-overexpressing flies, R8 axons stay anchored at the M1 layer (Fig.2C). From these results, we propose that Gogo recognizes a positive cue at the temporal layer M1. The protein Fmi has a very similar phenotype to Gogo in photoreceptor axon guidance as well as in dendrite formation. This led us test whether these two proteins collaborate. We first investigated the genetic interaction of Gogo and Fmi by overexpressing both proteins in the eye. Although overexpression of each protein separately only generates a mild rough eye phenotype, the eye is dramatically affected when both proteins are overexpressed. We also observed defects in the targeting of R7 axons: they frequently stopped at the M3 layer instead of M6 (Fig 3, compare A’ and B’ with C’). We further showed that Gogo and Fmi interact physically using two different assays. First, we used the fact that Fmi can generate homophilic interaction and accumulate at cell-cell contact in cultured cells. We co-transfected these cells with Gogo, and observed that Gogo co-accumulated with Fmi at cell-cell borders. We are now investigating how the interaction of Gogo and Fmi influence the targeting specificity of photoreceptor axons. Other projects in the lab include: |
|